
Cette chronique découle du courrier d’un lecteur. Elle traite d’un sujet sur lequel la religion de certains apiculteurs est déjà faite, même si les études sont peu nombreuses et les résultats contradictoires.
La question est donc sans réponse certaine. Cela arrive dans l’apidologie, d’autant plus que, pour ressasser une vieille formule, le vivant n’est pas une machine et nous n’avons pas toujours toutes les données pour le comprendre avec certitude. Chaque question que les chercheurs tentent de résoudre soulève parfois beaucoup de nouvelles interrogations.
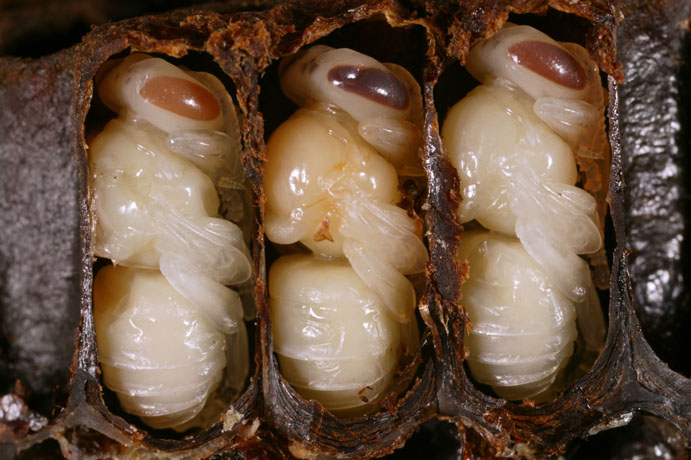
Un lecteur de la chronique du numéro 710, dans laquelle nous parlions des « fratries », a donc écrit : une question me taraude depuis longtemps : au moment de la fièvre d’essaimage ou de la supersédure, les larves choisies pour être élevées en reines sont-elles des super-sœurs ou sont-elles choisies au hasard ? Pour ma part, je pense qu’elles sont choisies dans le groupe de super-sœurs le plus nombreux, me référant à la loi de la nature qui avantage souvent le plus fort. Si la reine mère a été fécondée par exemple par 7 mâles A, 3 mâles B et 5 mâles C, je pense que les larves appelées à devenir reines seraient élevées dans le groupe A. A-t-on déjà, dans le monde scientifique, étudié et vérifié si les reines élevées naturellement étaient génétiquement des super-sœurs ou non ?
Essaimage et supersédure
Relisons la question soigneusement et pas à pas. Au départ, notre lecteur demande : au moment de la fièvre d’essaimage ou de la
supersédure, les larves choisies pour être élevées en reines sont-elles des super-sœurs ou sont-elles choisies au hasard ?
1. Si on veut vraiment être rigoureux, il pourrait y avoir dès les premiers mots plutôt deux questions qu’une seule. On y suppose en effet que l’essaimage et la supersédure sont un même environnement, qu’ils fonctionnent de la même façon, qu’ils sont deux processus assimilables l’un à l’autre. Est-ce forcément a priori la même situation ? Dans un cas, c’est la colonie le super-organisme abeille tout entier qui se multiplie (quasiment comme une amibe se dédouble), dans l’autre c’est seulement le remplacement par l’animal d’un de ses composants.
Les deux phénomènes n’arrivent pas tout à fait à la même période et dans les mêmes conditions, l’un (la supersédure) suit l’autre (l’essaimage) assez
souvent mais il ne semble jamais le précéder, du moins sous nos climats et d’après ce que nous croyons savoir.
Cela dit, chez certains cas, il paraîtrait que la nouvelle reine et l’ancienne peuvent coexister plus souvent dans la ruche avant l’essaimage, une situation qu’avait utilisé Patrice Serres dans Le Bal des abeilles, sur les indications de Rémy Chauvin. Si nous rentrons nous perdre dans le fond des détails, peut-on dire que le nombre des cellules royales et leurs emplacements habituels sont assimilables dans les deux procédés ?
J’ai cru un temps innocemment que les emplacements des cellules royales (et donc les procédures) étaient un peu différentes (peut-être plus souvent au centre du couvain dans un cas, peut-être plus souvent au bas d’un rayon dans l’autre), mais des gens plus expérimentés que moi m’ont bien expliqué
qu’on n’en savait rien du tout, que tous les cas étaient possibles et qu’il n’y avait pas de règle, fut-elle statistique. Ainsi pour les reines de sauveté, on sait que des tas de cellules royales seront construites aussi sur les bords de cadres puis souvent (mais pas toujours) abandonnées au profit de celles qui sont dans la zone du couvain, probablement pour de simples raisons de température, afin de garantir un meilleur développement de la future reine. On peut ici supposer que le critère de choix est bien plus le bon développement de la future reine que ses origines patrilinéaires.
Des deux procédures de l’essaimage et de la supersédure, il faudrait peut-être aussi vraiment vérifier que, dans les fréquences statistiques de leur occurrence selon les races, ils s’excluent l’un l’autre et que les races, voire les peuples, les souches, qui essaiment beaucoup restent celles ou ceux qui font peu de supersédure (et inversement). C’est une idée qui semble logique a priori mais qui ne prouve pas non plus que les deux systèmes sont assimilables l’un à l’autre. A-t-on également tenté de mesurer l’occurrence de la supersédure dans les essaims enruchés après l’essaimage ?
Bref, il vaudrait peut-être mieux poser la question ainsi : au moment de la fièvre d’essaimage, les larves choisies pour être élevées en reines sont-elles des super-sœurs ou sont-elles choisies au hasard ? Même question au moment de la supersédure.
2. Pour continuer cette rigueur, il faudrait d’ailleurs se rappeler qu’il existe un troisième cas de sélection de larves (ou plutôt d’œufs) devant donner des reines : la procédure d’urgence de la reine de sauveté.
On devrait donc poser la question et étudier la chose dans trois environnements semblables mais pas forcément identiques.
Comment choisir le « bon » spermatozoïde ?
La question continue par : « les larves choisies pour être élevées en reines » et on bute ici forcément sur le mot « choisies ». Sont-elles choisies ? Par qui seraient-elles choisies, ou par quel mécanisme, et à quel moment ?
Par la reine ? Nous savons que la reine peut choisir de féconder un œuf ou de ne pas le féconder, c’est-à-dire de faire un mâle ou une femelle. Nous savons que, avant de pondre, elle mesure les dimensions de la cellule et qu’elle sait donc s’il s’agit-là d’une cellule de mâle ou de femelle. Mais que peut-elle au- delà ? Comment pourrait-elle choisir un spermatozoïde parmi d’autres ? Elle n’a pas les moyens de les repérer et d’extirper tel ou tel parmi les milliers qui peuplent sa spermathèque. Elle doit pondre et laisser à ses ouvrières le soin de détruire ceux des œufs qui seraient imparfaits ou qui proviendraient d’ouvrières rebelles. Elle est souvent dans un rythme très soutenu pour pondre et les œufs déboulent parfois de son utérus les uns après les autres. Alors ? Car c’est au stade de l’œuf, ou plutôt de la cellule dans laquelle elle va le pondre, que s’effectue le « choix » du spermatozoïde. On peut légitimement craindre que ce ne soit pas un vrai choix.
Comment discriminer entre les œufs ?
Restent maintenant les cellules royales que les abeilles vont édifier. On dit souvent que ces cellules sont construites à partir d’une alvéole contenant déjà un œuf fécondé récemment pondu. Peut-on assimiler cela au cas des reines de sauveté pour lesquelles les ouvrières improvisent à partir d’œufs déjà en place dans un couvain préexistant ? Que les ouvrières choisissent, c’est vraisemblable et elles ne prendront sans doute pas un œuf malade ou imparfait. Mais peuvent elles discriminer au-delà ? Normalement, non. Donc pas de choix possible. Peuvent-elles reconnaître le père d’un œuf fécondé ? Dans l’état actuel de nos connaissances, ce n’est pas certain du tout.
Népotisme ? Possible ou impossible ?
La question devient ensuite : « Pour ma part, je pense qu’elles sont choisies dans le groupe de super-sœurs le plus nombreux, me référant à la loi de la nature qui avantage souvent le plus fort.
Si la reine mère a été fécondée par exemple par 7 mâles A, 3 mâles B et 5 mâles C, je pense que les larves appelées à devenir reines seraient élevées dans le groupe A.
A-t-on déjà, dans le monde scientifique, étudié et vérifié si les reines élevées naturellement étaient génétiquement des super-sœurs ou non ? »
On pourrait suivre l’assimilation faite entre le groupe le plus nombreux et le groupe le plus fort. Cela dit, la « loi de la nature qui avantage souvent le plus fort » sonne comme un néo-darwinisme un peu hâtif. Il vaudrait mieux parler du plus apte et non du plus fort. Ensuite la théorie darwinienne, comme ses prolongements modernes en milieu scientifique sont tout de même très nuancés.
Cela dit, les plus nombreux, même s’ils ne sont pas les plus forts, devraient logiquement avoir plus de chance dans la course à la royauté. S’il y a vraiment un groupe sensiblement plus nombreux et une large diversité dans les spermatozoïdes et dans la période de ponte, ce groupe le plus nombreux aurait-il donc mathématiquement plus de chances d’avoir des descendants dans les œufs qui seront fécondés ? Mouais, peut-être mais sur quelle période et sur combien d’années ?
Répartition dans la spermathèque et dans la ponte
Pourquoi pas mais… On ne sait toujours pas vraiment comment fonctionne la spermathèque : y a-t-il des séquences homogènes de spermatozoïdes dans la ponte de la reine à partir de regroupements, d’amas homogènes, dans la spermathèque ? Probablement oui, puisque la reine a été fécondée par beaucoup de faux bourdons à la suite. Chacun d’eux meurt immédiatement après la copulation et tombe sur le sol en laissant dans l’appareil génital de la reine de 6 à 12 millions de spermatozoïdes, ainsi qu’un « bouchon d’accouplement », c’est-à-dire essentiellement l’endophallus qui lui a été arraché mais aussi une sécrétion particulière qui sert sans doute à empêcher la perte des spermatozoïdes.
Le mâle suivant qui se présente pour la copulation va expulser cette barrière et procéder à une nouvelle copulation. L’opération complète dure 2 secondes seulement ! Le dernier endophallus arraché aux mâles (les « voiles de la mariée », disaient les vieux mouchiers) sera retiré par les ouvrières au retour de la reine à la colonie. La plus grande partie du sperme accumulé dans les oviductes va être expulsée (95 % du sperme), mais 5,5 millions environ de spermatozoïdes migreront lentement vers la spermathèque. Le processus complet durera tout de même une quarantaine d’heures ! On peut penser que le brassage final dans la spermathèque ne sera pas bien élaboré, que les spermatozoïdes d’un même mâle vont migrer plus ou moins ensemble vers la spermathèque, qu’ils y vivront ensemble avant d’être utilisés ensemble. C’est peut-être ce qui crée des castes particulières dans la colonie à certaines époques, et non un népotisme qu’on n’a guère pu prouver, ni expliquer, même s’il a été constaté statistiquement.
Cela dit, la reine qui pond dans des cellules royales déjà prêtes (ça existe) ou bien la reine qui pond dans des cellules normales que les ouvrières transformeront ensuite en cellules royales (ça existe aussi) n’est jamais forcée de tout faire en série. Ce qui veut dire que ses pontes « royales » ne se suivent pas forcément. D’ailleurs, au moins dans le cas de l’essaimage, entre deux cellules royales édifiées par les ouvrières, il y aura parfois plusieurs jours d’intervalle. Pour couronner le tout, les ouvrières mettront au départ en chantier beaucoup plus de cellules royales qu’il n’y en aura de terminées et de remplies à l’arrivée, et dans des coins parfois bien éloignés. Aussi notre séquence, cette idée d’une suite d’œufs fécondés par un même mâle et conséquemment pondus à la suite, a déjà du plomb dans l’aile. Plus il y a eu de mâles différents pour participer à la fécondation et plus elle sera d’ailleurs moins probable.
D’ailleurs on a aussi noté, dans le cas des destructions de cellules royales de reines de sauveté par les ouvrières, que ces destructions avaient lieu dans les zones où le couvain adjacent était le plus vieux. La décision de détruire cette cellule pourrait donc bien correspondre simplement à une préférence pour les œufs les plus frais qui donneront les meilleures reines, non pas à un népotisme quelconque.
On peut seulement penser qu’il y a plus de chance pour les plus nombreux de donner une nouvelle reine. C’est pourtant une probabilité bien faible, si on veut bien considérer qu’elle concerne déjà une bonne quinzaine de mâles et qu’elle s’étend, si on veut rester rigoureux, sur environ 5 ans, durée biologique moyenne de vie d’une reine et de la vidange théorique de l’intégralité de sa spermathèque.
En outre, les plus nombreux sont-ils forcément les plus aptes ? J’aurai tout de même un peu de méfiance et de réticence car il peut et il doit exister des cas, surtout à notre époque, où le choix des mâles est limité et la répartition en est déséquilibrée tout simplement parce qu’il n’y a pas assez de ruches proches (et encore moins de colonies sauvages proches) pour alimenter la congrégation de mâles qui ira au devant de la reine. Dans ce cas, la reine déjà fécondée le sera avec moins de diversité. Si la quantité des mâles disponibles s’en ressent et que cela fait que la reine est insuffisamment fécondée, elle refera un second (voire un troisième) vol de fécondation dans les jours à venir. Mais trouvera-t-elle vraiment une autre répartition des mâles disponibles lors de ces nouveaux vols ? Le déséquilibre persistera alors que, nous a-t-on appris, il est nécessaire qu’il y ait le maximum de diversité. Et donc un maximum aussi de reines d’origines différentes et de bourdons qui les ont fécondées elles-mêmes. (Si la situation dure trop longtemps et sur plusieurs générations, on pourrait même aboutir à terme à une certaine fragilité des souches. Le petit nombre d’éleveurs de reines aux USA et leurs déséquilibres fait d’ailleurs également partie des sujets étudiés dans le cadre des multiples recherches initiées par le CDD, le syndrome d’effondrement des colonies).
Pour la même raison que précédemment, pourquoi et comment pourrait-on (« on » étant à la rigueur la mère et/ou surtout les nourrices) sélectionner les larves à partir d’un œuf ? On ne voit pas.
Rien ne permet donc de présupposer avec fermeté qu’il y aurait des super-sisters dans les œufs qui remplissent les cellules royales, et c’est pourquoi, à ma connaissance, la chose n’a pas été étudiée. Citons aussi les études faites sur les croisements d’abeilles africaines et européennes (les fameuses abeilles tueuses). On a constaté que, dans une colonie métissée (africanisée), les reines d’origine plus africaine que les autres, de pères donc plus africains que les autres, avaient une gestation un tout petit peu plus courte. Elles éclosent donc avant les autres et cet avantage leur permet de tuer plus facilement les autres futures reines. Si bien que les colonies métisses se ré-africanisent peu à peu et finissent doucement par retourner à une seule des origines, malheureusement la moins souhaitable. Mais, a contrario, on sait donc qu’il y a plusieurs pères pour les reines qui éclosent, ce que la formulation de notre question ne conteste d’ailleurs pas.
Reste le cas des reines de sauveté pour lesquelles les ouvrières vont (généralement ?) coller un œuf dans une goutte de gelée royale au fond d’une cellule qu’elles vont construire (mais parfois aussi qu’elles viennent de construire ?). Comment ont-elles choisi cet œuf ?
Les théories de l’altruisme privilégient une signature de père commun, mais on ne voit pas ici comment cela serait possible. D’ailleurs, les études pour le rechercher et le prouver ont échoué. Si on a deux lignages seulement, trois lignages à la rigueur, dans des colonies artificielles évidemment (ou bien aussi dans des ruchers peu importants et isolés, sur une longue période ?), on croit constater une certaine préférence, un certain népotisme. Ainsi, dans la trophallaxie : les donneuses privilégieraient leurs super-sœurs. Cependant, si on en a plus de trois lignages, il n’y a plus aucune reconnaissance des lignages. C’est en conformité avec ce qu’on sait des hydrocarbones cuticulaires auxquels nous avions déjà consacré deux chroniques (Abeilles et Fleurs n° 701 et n° 702).
Trichage et sabotage dans la course à l’éclosion ?
Pourtant, il reste encore une possibilité de népotisme dans la sélection de la reine finale. Mais nous entrons presque dans le roman policier ou sociologique.
Nous avons vu que l’éclosion des fratries africaines leur donne un avantage certain dans un peuple métissé d’abeilles africanisées.
Peut-on imaginer qu’à cet avantage naturel soit artificiellement substitué un handicap exogène ? Peut-on imaginer des jeunes reines, prêtes à éclore, et
des ouvrières malintentionnées qui les empêcheraient de sortir de leurs cellules royales en premier, dans le seul but de favoriser une supersister encore dans sa cellule, une supersister qu’elles ont pu percevoir et qu’elles veulent avantager ? Certains chercheurs ont envisagé cette possibilité, avec des résultats divers. Nous en parlerons dans une prochaine chronique.
Article paru dans la revue Abeilles & Fleurs N° 713 – Février 2010